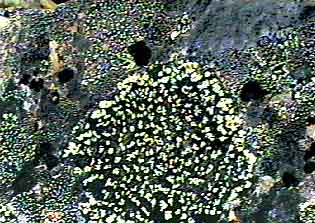
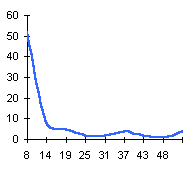
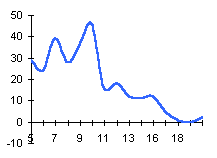
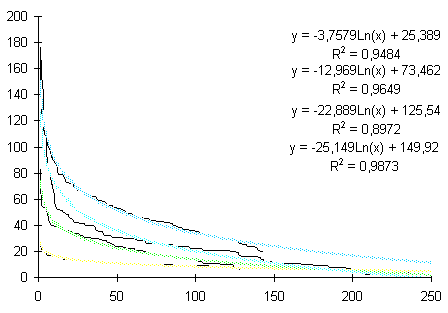
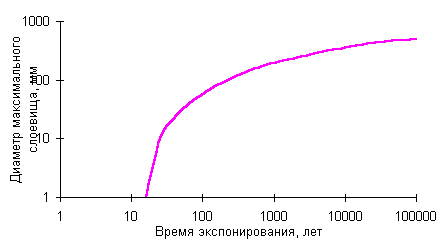
лишайника-индикатора, а, b и с - коэффициенты,
зависящие от особенностей используемого вида и физико-климатических условий
района датирования. Очевидно, что для нахождения этих коэффициентов
необходимо решить систему из трех уравнений. Для территории Норвегии этим
исследователем коэффициенты а, b и с
были оценены в 1,8143, 0,0062 и 50 соответственно. В предлагаемой
автором зависимости не возраст, а логарифм возраста линейно связан с диаметром
максимального лишайника. Выполненный нами анализ данной зависимости при
разных значениях x показал, что она отражает представления
Р.Е. Бешеля о фазе быстрого нелинейного роста и, следующей за ней линейной
фазе. Однако, по данной функции в интервале возрастов более 100 лет, она
принимает форму, близкую к линейной. Что противоречит условию об предельных
размерах роста слоевищ накипных лишайников. А.А.Никоновым с соавторами
для построения вероятностной модели роста локальных популяций лишайника
индикатора использовался известный закон замедления роста численности живых
организмов - Т.Р. Мальтуса. Профессионально математически постороенная
авторами модель весьма сложна для практического использования, к тому же
в ее основе приняты некоторые априорные допущения, противоречащие
законам конкуренции и популяционно-ценотических отношений в локальных сообществах
накипных лишайников [Галанин, 1997; 1999]. Тем не менее, среди известных
лихенометрических методик она является наиболее удовлетворительной,
позволяя рассчитать ошибки возрастных оценок.
В качестве замедляющей функции нами был
принята убывающая экспонента и допущение, что замедление прироста максимального
теоретического диаметра локальной выборки происходит по экспоненциальному
закону. Пусть в первый год после заселения какой-либо “свежей” каменной
поверхности прирост используемой статистики (максимального теоретического
диаметра) лишайников-индикаторов составил ао
мм/год.
Тогда средний выборочный прирост аi
в год i с учетом экспоненциального замедления будет: ai=
aо
* e^ (-i / f), где f
- некий параметр замедления. Максимальный теоретический диаметр dt
на поверхности возраста t лет будет равен сумме
годовых приростов:
-
t
-
dt =a0*
Se^
(-i / f)
,
-
i= 1
а после интегрирования
t
dt »a0*тe^
(-i / f)(di) = a0f*(e^
(-1 / f) - e^ (-i / f))
.
1
Величина e^
(-1 / f) уже при
f >>10
приближается к 1, следовательно, для приближенных
расчетов можно использовать формулу:
d »a0f
* (1 - e^ (-i / f))(1)
.
На поверхности с небольшим временем
экспонирования замедление роста слоевищ существенно не проявится,
поэтому:
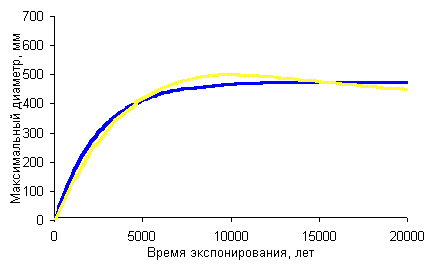
dt »a0t (2).
Можно показать, что при большом возрасте
каменной поверхности (t ®Ґ)
величина e^
- t/f ® 0,
следовательно, в этих случаях будет верна формула
dt ®Ґ
» a0 f (3),
где dt®Ґ
- средний выборочный диаметр на поверхности большого (условно бесконечного)
времени экспонирования.
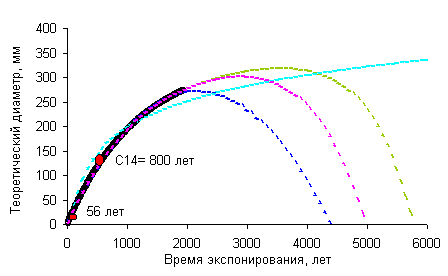
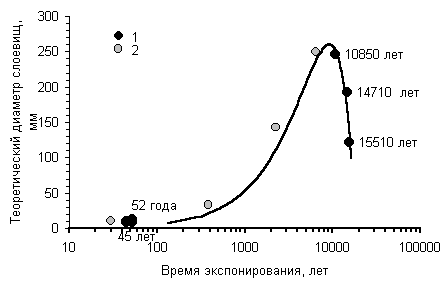
Предлагаемая методика позволяет построить
приближенную кривую роста для района, в котором имеется 1 или более независимо
датированных поверхностей колонизованных накипными лишайниками. Начальный
годовой прирост приближенно вычисляется по формуле 2,
используя результаты измерений талломов на поверхности известного возраста,
например горной выработке, карьере, с использованием дендрохронологических
данных и др. Параметр замедления роста рассчитывается, используя приближение
3,
по результатам изменений диаметров слоевищ на поверхности древнего условно
бесконечного возраста. Такие поверхности хорошо определяются по степени
выветрелости обломков, характеру склоновых процессов, лишайниковым сукцессиям
и другим геоморфологическим признакам.
Параметры роста в разных районах неодинаковы
и зависят от их физико-географических условий, определяющих продолжительность
вегетационного периода лишайников. Для субарктической тундры побережья
Анадырского лимана параметры ао
и f для среднего значения ста максимальных особей соответственно
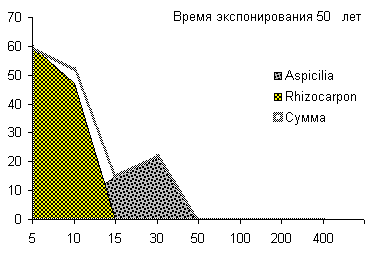
составляют: секция Rhizocarpon - 0,14±0,01 мм/год и 526±95 лет,
род Aspicilia - 0,15±0,01 мм/год и 733±115 лет. В лесотундровой
зоне центральной части Корякского нагорья для секции Rhizocarpon - 0,18±0,03
мм/год и 452±88 лет, а начальный годовой прирост листоватого лишайника
Parmelia sp. - 0,47±0,08 мм/год. В Примагаданье (зона северной тайги)
параметры роста лишайников рода Rhizocarpon секции Rhizocarpon
составляют 0,23±0,03 мм/год и 419±86 лет. Таким образом, скорости
роста исследуемых лишайников в разных физико-географических районах существенно
различаются. Наиболее низкие годовые приросты и, соответственно, более
медленный рост и развитие популяций накипных лишайников характерно для
побережья Анадырского лимана, что не противоречит условиям субарктического
климата. Параметры роста накипных лишайников в Северной Корякии и Примагаданье
близки по значениям, что, по-видимому, можно объяснить близкими климатическими
характеристиками этих районов. |